FROM: Scientific American ~ February, 2006
By David A. Sinclair and Lenny Guarente
A compound found in red wine may extend the human life span.
A report from the front lines.
A handful of genes that control the body's defenses during hard times can also dramatically improve health and prolong life in diverse organisms. Understanding how they work may reveal the keys to extending human life span while banishing diseases of old age
TAPPING THE POWER of longevity genes could change the arc of a typical human lifetime: instead of vitality and growth giving way to the decline of old age, a person might be able to retain the youthfulness he feels at 50 when he is 70, 90 or well past 100.
You can assume quite a bit about the state of a used car just from its mileage and model year. The wear and tear of heavy driving and the passage of time will have taken an inevitable toll. The same appears to be true of aging in people, but the analogy is flawed because of a crucial difference between inanimate machines and living creatures: deterioration is not inexorable in biological systems, which can respond to their environments and use their own energy to defend and repair themselves.
At one time, scientists believed aging to be not just deterioration but an active continuation of an organism's genetically programmed development. Once an individual achieved maturity, "aging genes" began to direct its progress toward the grave. This idea has been discredited, and conventional wisdom now holds that aging really is just wearing out over time because the body's normal maintenance and repair mechanisms simply wane. Evolutionary natural selection, the logic goes, has no reason to keep them working once an organism has passed its reproductive age.
Yet we and other researchers have found that a family of genes involved in an organism's ability to withstand a stressful environment, such as excessive heat or scarcity of food or water, have the power to keep its natural defense and repair activities going strong regardless of age. By optimizing the body's functioning for survival, these genes maximize the individual's chances of getting through the crisis. And if they remain activated long enough, they can also dramatically enhance the organism's health and extend its life span. In essence, they represent the opposite of aging genes--longevity genes.
We began investigating this idea nearly 15 years ago by imagining that evolution would have favored a universal regulatory system to coordinate this well-known response to environmental stress. If we could identify the gene or genes that serve as its master controllers and thereby act as master regulators of an organism's life span, these natural defense mechanisms might be turned into weapons against the diseases and decline that are now apparently synonymous with human aging.
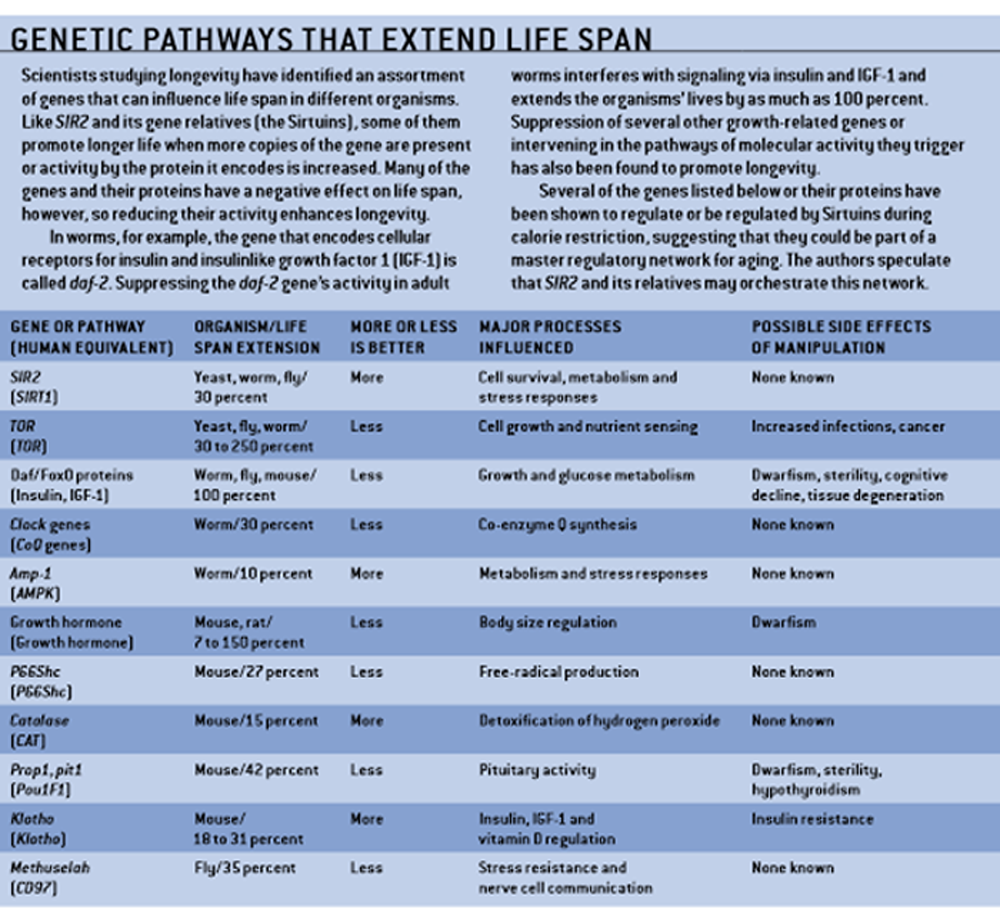
Many recently discovered genes, known by such cryptic names as daf-2, pit-1, amp-1, clk-1 and p66Shc, have been found to affect stress resistance and life span in laboratory organisms, suggesting that they could be part of a fundamental mechanism for surviving adversity. But our own two laboratories have focused on a gene called SIR2, variants of which are present in all organisms studied so far, from yeast to humans. Extra copies of the gene increase longevity in creatures as diverse as yeast, roundworms and fruit flies, and we are working to determine whether it does the same for larger animals, such as mice.
As one of the first longevity genes to have been identified, SIR2 is the best characterized, so we will focus here on its workings. They illustrate how a genetically regulated survival mechanism can extend life and improve health, and growing evidence suggests that SIR2 may be the key regulator of that mechanism.
Silence Is Golden
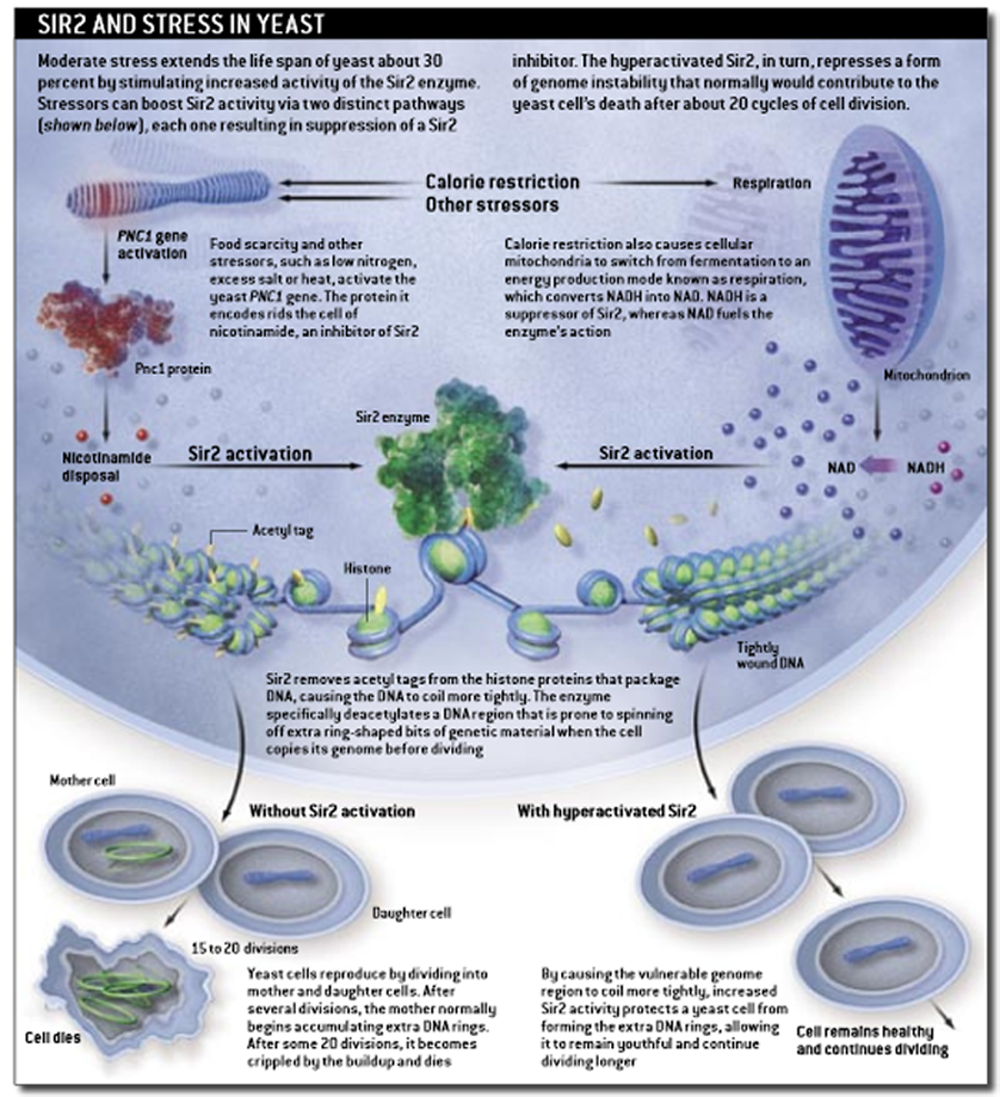
We first discovered that SIR2 is a longevity gene by asking what causes individual baker's yeast cells to grow old and whether a single gene might control aging in this simple organism. The notion that an understanding of yeast life span would tell us anything about human aging was deemed preposterous by many. Aging in yeast is measured by counting how many times mother cells divide to produce daughters before dying. A typical yeast cell's life span is about 20 divisions.
One of us (Guarente) began by screening yeast colonies for unusually long-lived cells in the hope of finding genes responsible for their longevity. This screen yielded a single mutation in a gene called SIR4, which encodes part of a complex of proteins containing the Sir2 enzyme. The mutation in SIR4 caused the Sir2 protein to gather at the most highly repetitive region of the yeast genome, a stretch containing the genes that encode the protein factories of the cell, known as ribosomal DNA (rDNA). More than 100 of these rDNA repeats exist in the average yeast cell's genome, and they are difficult to maintain in a stable state. Repetitive sequences are prone to "recombining" with one another, a process that in humans can lead to numerous illnesses, such as cancer and Huntington's disease. Our yeast findings suggested that aging in mother cells was caused by some form of rDNA instability that was mitigated by the Sir proteins.
In fact, we found a surprising kind of rDNA instability. After dividing several times, yeast mother cells spin off extra copies of the rDNA as circular rings that pop out of the genome. These extrachromosomal rDNA circles (ERCs) are copied along with the mother cell's chromosomes prior to cell division but remain in the mother cell's nucleus afterward. Thus, a mother cell accumulates an ever increasing number of circles that eventually spell her doom, possibly because copying the ERCs consumes so many resources that she can no longer manage to replicate her own genome.
When an extra copy of the SIR2 gene was added to the yeast cell, however, formation of the rDNA circles was repressed and the cell's life span was extended by 30 percent. That finding explained how sir2 could act as a longevity gene in yeast, but amazingly, we soon discovered that extra copies of the sir2 gene also extended the life span of roundworms by as much as 50 percent. We were surprised not only by this commonality in organisms separated by a vast evolutionary distance but by the fact that the adult worm body contains only nondividing cells--thus, the replicative aging mechanism in yeast could not apply to worms. We wanted to know exactly what the SIR2 gene does.
As we soon discovered, the gene encodes an enzyme with a completely novel activity. Cellular DNA is wrapped around a complex of packaging proteins called histones. These bear chemical tags, such as acetyl groups, that determine how snugly the histones package DNA. Removing acetyl groups from histones tightens the packaging further and renders the DNA inaccessible to the enzymes responsible for popping the rDNA circles out of the chromosome. This deacetylated form of DNA is said to be silent because any genes in these regions of the genome are rendered inaccessible to being activated.
Sir proteins were already known to be involved in gene silencing--indeed, SIR stands for silent information regulator. Sir2 is one of several enzymes that remove acetyl tags from the histones, but we discovered that it is unique in that its enzymatic activity absolutely requires a ubiquitous small molecule called NAD, which has long been known as a conduit of many metabolic reactions in cells. This association between Sir2 and NAD was exciting because it linked Sir2 activity to metabolism and thus potentially to the relation between diet and aging observed in calorie restriction.
The Calorie Connection
Restricting an animal's calorie intake is the most famous intervention known to extend life span. Discovered more than 70 years ago, it is still the only one absolutely proven to work. The restricted regime typically involves reducing an individual's food consumption by 30 to 40 percent compared with what is considered normal for its species. Animals ranging from rats and mice to dogs and possibly primates that remain on this diet not only live longer but are far healthier during their prolonged lives. Most diseases, including cancer, diabetes and even neurodegenerative illnesses, are forestalled. The organism seems to be supercharged for survival. The only apparent trade-off in some creatures is a loss of fertility.
Understanding the mechanisms by which calorie restriction works and developing medicines that reproduce its health benefits have been tantalizing goals for decades [see "The Serious Search for an Antiaging Pill," by Mark A. Lane, Donald K. Ingram and George S. Roth; Scientific American: The Science of Staying Young, 2004]. The phenomenon was long attributed to a simple slowing down of metabolism--cells' production of energy from fuel molecules--and therefore reduction of its toxic by-products in response to less food.
But this view now appears to be incorrect. Calorie restriction does not slow metabolism in mammals, and in yeast and worms, metabolism is both sped up and altered by the diet. We believe, therefore, that calorie restriction is a biological stressor like natural food scarcity that induces a defensive response to boost the organism's chances of survival. In mammals, its effects include changes in cellular defenses, repair, energy production and activation of programmed cell death known as apoptosis. We were eager to know what part Sir2 might play in such changes, so we looked first at its role during calorie restriction in simple organisms
In yeast, we have found that restricting food availability affects two pathways that increase Sir2 enzymatic activity in the cells. On one hand, calorie restriction turns on a gene called PNC1, which produces an enzyme that rids cells of nicotinamide, a small molecule similar to vitamin B3 that normally represses Sir2. Consistent with the idea that calorie restriction is a stressor that activates a survival response, PNC1 is also stimulated by other mild stressors known to extend yeast life span, such as increased temperature or excessive amounts of salt.
A second pathway induced in yeast by restricted calories is respiration, a mode of energy production that creates NAD as a by-product while lowering levels of its counterpart, NADH. It turns out that not only does NAD activate Sir2, but NADH is an inhibitor of the enzyme, so altering the cell's NAD/NADH ratio profoundly influences Sir2 activity.
Having seen how life-extending biological stress increases Sir2 activity, the question became, Is Sir2 necessary to produce the longevity? The answer appears to be a resounding "yes." One way to test whether Sir2 is essential to this process is to remove its gene and determine whether the effect remains. In organisms as complex as fruit flies, calorie restriction does require SIR2 to extend life span. And because the body of an adult fruit fly contains numerous tissues that are analogous to mammalian organs, we suspect that calorie restriction in mammals is also likely to require SIR2.
Yet if humans are ever to reap the health benefits of calorie restriction, radical dieting is not a reasonable option. Drugs that can modulate the activity of Sir2 and its siblings (collectively referred to as Sirtuins) in a similar manner will be needed. Just such a Sirtuin-activating compound, or STAC, called resveratrol has proven particularly interesting. Resveratrol is a small molecule present in red wine and manufactured by a variety of plants when they are stressed. At least 18 other compounds produced by plants in response to stress have also been found to modulate Sirtuins, suggest?-ing that the plants may use such mole?-cules to control their own Sir2 enzymes.
Feeding resveratrol to yeast, worms or flies or placing them on a calorie-restricted diet extends their life spans about 30 percent, but only if they possess the SIR2 gene. Moreover, a fly that overproduces Sir2 has an increased life span that cannot be further extended by resveratrol or calorie restriction. The simplest interpretation is that calorie restriction and resveratrol each prolong the lives of fruit flies by activating Sir2.
Resveratrol-fed flies not only live longer, despite eating as much as they want, but they do not suffer from the reduced fertility often caused by calorie restriction. This is welcome news for those of us hoping to treat human diseases with molecules that target Sir2 enzymes. But first we want a better understanding of the role of Sir2 in mammals.
Leader of the Band
The mammalian version of the yeast SIR2 gene is known as SIRT1 ("SIR2 homolog 1"). It encodes a protein, Sirt1, that has the same enzymatic activity as Sir2 but that also deacetylates a wider variety of proteins both inside the cell nucleus and out in the cellular cytoplasm. Several of these proteins targeted by Sirt1 have been identified and are known to control critical processes, including apoptosis, cell defenses and metabolism. The potential longevity-enhancing role of the SIR2 gene family seems, therefore, to be preserved in mammals. But not surprisingly in larger and more complex organisms, the pathways by which Sirtuins achieve their effect have grown considerably more complicated as well.
 |
Increased Sirt1 in mice and rats, for example, allows some of the animals' cells to survive in the face of stress that would normally trigger their programmed suicide. Sirt1 does this by regulating the activity of several other key cellular proteins, such as p53, FoxO and Ku70, that are involved either in setting a threshold for apoptosis or in prompting cell repair. Sirt1 thus enhances cellular repair mechanisms while buying time for them to work.
Over the course of a lifetime, cell loss from apoptosis may be an important factor in aging, particularly in nonrenewable tissues such as the heart and brain, and slowing cell death may be one way Sirtuins promote health and longevity. A striking example of Sirt1's ability to foster survival in mammalian cells can be seen in the Wallerian mutant strain of mouse. In these mice, a single gene is duplicated, and the mutation renders their neurons highly resistant to stress, which protects them against stroke, chemotherapy-induced toxicity and neurodegenerative diseases.
In 2004 Jeffrey D. Milbrandt of Washington University in St. Louis and his colleagues showed that the Wallerian gene mutation in these mice increases the activity of an enzyme that makes NAD, and the additional NAD appears to protect the neurons by activating Sirt1. Moreover, Milbrandt's group found that STACs such as resveratrol conferred a protective effect on the neurons of normal mice similar to the Wallerian mutation.
In a more recent study by Christian N??? of the French National Institute of Health and Medical Research, resveratrol and another STAC, fisetin, were shown to prevent nerve cells from dying in two different animal models (worm and mouse) of human Huntington's disease. In both cases, the protection by STACs required Sirtuin gene activity.
The protective effect of Sirtuins in individual cells is becoming increasingly clear. But if these genes are the mediators of calorie restriction's benefits, an unsolved puzzle remains how diet can regulate their activities and thus the rate of aging in an entire animal. Recent research by Pere Puigserver of the Johns Hopkins University School of Medicine and his colleagues has shown that NAD levels rise in liver cells under fasting conditions, prompting increased Sirt1 activity. Among the proteins Sirt1 acts on is an important regulator of gene transcription called PGC-1, which then causes changes in the cell's glucose metabolism. Thus, Sirt1 was found to act both as a sensor of nutrient availability and a regulator of the liver's response.
Similar data have given rise to the idea that Sirt1 is a central metabolic regulator in liver, muscle and fat cells because it senses dietary variations via changes in the NAD/NADH ratio within cells and then exerts far-reaching effects on the pattern of gene transcription in those tissues. This model would explain how Sirt1 may integrate many of the genes and pathways that affect longevity described on page 54.
 |
More than one mechanism may mediate Sirt1's bodywide activities, however. Another appealing hypothesis is that mammals register their food availability by the amount of energy they have stored in the form of body fat. Fat cells secrete hormones that convey signals to the other tissues in the body, but their message depends on the levels of fat stored. By reducing fat stores, calorie restriction may establish a pattern of hormone signals that communicates "scarcity," which activates cell defenses. Consistent with this idea is the fact that mice genetically engineered to be extra lean regardless of their food intake tend to live longer.
This possibility led us to wonder whether Sirt1, in turn, also regulates fat storage in response to diet. Indeed, Sirt1 activity is increased in fat cells after food limitation, causing fat stores to move from the cells into the bloodstream for conversion to energy in other tissues. We surmise that Sirt1 senses the diet, then dictates the level of fat storage and thus the pattern of hormones produced by fat cells. This effect on fat and the signals it sends would, in turn, set the pace of aging in the entire organism and make Sirt1 a key regulator of the longevity conferred by calorie restriction in mammals. It would also closely link aging and metabolic diseases, including type 2 diabetes, associated with excess fat. Intervening pharmacologically in the Sirt1 pathway in fat cells might therefore forestall not only aging but also specific ailments.
Another critical process modified by Sirt1 is inflammation, which is involved in a number of disorders, including cancer, arthritis, asthma, heart disease and neurodegeneration. Recent work by Martin W. Mayo and his colleagues at the University of Virginia has shown that Sirt1 inhibits NF-B, a protein complex that promotes the inflammatory response. The Sirt1-activating compound resveratrol has the same effect. This finding is particularly encouraging, both because the search for molecules that inhibit NF-B is a highly active area of drug development and because another well-known effect of calorie restriction is its ability to suppress excessive inflammation.
If SIR2 is thus the master controller of a regulatory system for aging that is activated by stress, it may function by acting as the conductor of an orchestra of players that includes hormonal networks, intracellular regulatory proteins and other genes associated with longevity. One of the more notable discoveries in recent years was that Sirt1 regulates production of insulin and insulinlike growth factor 1 (IGF-1) and that those two powerful signaling molecules, in turn, seem to regulate Sirt1 production as part of a complex feedback loop. The relation between Sirt1, IGF-1 and insulin is intriguing because it explains how Sirt1 activity in one tissue might be communicated to other cells in the body. Moreover, circulating levels of insulin and IGF-1 are known to dictate life span in various organisms--worms, flies, mice, possibly ourselves.
From Defense to Advance
Because people have sought to slow aging for tens of thousands of years without success, some may find it hard to accept that human aging might be controlled by tweaking a handful of genes. Yet we know it is possible to forestall aging in mammals with a simple dietary change: calorie restriction works. And we have shown that Sirtuin genes control many of the same molecular pathways as calorie restriction. Without actually knowing the precise, and potentially myriad, causes of aging, we have already demonstrated in a variety of life-forms that it can be delayed by manipulating a few regulators and letting them take care of the organisms' health.
We also know that the SIR2 family of genes evolved far back in time because today they are found in organisms ranging from baker's yeast, Leishmania parasites and roundworms to flies and humans. In all these organisms but the last, which has not yet been tested, Sirtuins dictate length of life. This fact alone convinces us that human Sirtuin genes probably hold the key to our health and longevity as well.
Both our labs are running carefully controlled mouse experiments that should soon tell us whether the SIRT1 gene controls health and life span in a mammal. We will not know definitively how Sirtuin genes affect human longevity for decades. Those who are hoping to pop a pill and live to 130 may have therefore been born a bit too early. Nevertheless, those of us already alive could live to see medications that modulate the activity of Sirtuin enzymes employed to treat specific conditions such as Alzheimer's, cancer, diabetes and heart disease. In fact, several such drugs have begun clinical trials for treatment of diabetes, herpes and neurodegenerative diseases.
And in the longer term, we expect that unlocking the secrets of longevity genes will allow society to go beyond treating illnesses associated with aging and prevent them from arising in the first place. It may seem hard to imagine what life will be like when people are able to feel youthful and live relatively free of today's diseases well into their 90s. Some may wonder whether tinkering with human life span is even a good idea. But at the beginning of the 20th century, life expectancy at birth was around 45 years. It has risen to about 75 thanks to the advent of antibiotics and public health measures that allow people to survive or avoid infectious diseases. Society adapted to that dramatic change in average longevity, and few people would want to return to life without those advances. No doubt, future generations accustomed to living past 100 will also look back at our current approaches to improving health as primitive relics of a bygone era.






