

A Complex Dietary Supplement Augments Spatial Learning,
Brain Mass, and Mitochondrial Electron Transport
Chain Activity in Aging MiceThis section is compiled by Frank M. Painter, D.C.
Send all comments or additions to: Frankp@chiro.org


FROM: Age (Dordr). 2013 (Feb); 35 (1): 23–33 ~ FULL TEXT
Vadim Aksenov, Jiangang Long, Jiankang Liu, Henry Szechtman, Parul Khanna, Sarthak Matravadia, and C. David Rollo
Department of Biology,
McMaster University,
1280 Main St W.,
Hamilton, ON, Canada.
We developed a complex dietary supplement designed to offset five key mechanisms of aging and tested its effectiveness in ameliorating age-related cognitive decline using a visually cued Morris water maze test. All younger mice (<1 year old) learned the task well. However, older untreated mice (>1 year) were unable to learn the maze even after 5 days, indicative of strong cognitive decline at older ages. In contrast, no cognitive decline was evident in older supplemented mice, even when ?2 years old. Supplemented older mice were nearly 50% better at locating the platform than age-matched controls. Brain weights of supplemented mice were significantly greater than controls, even at younger ages. Reversal of cognitive decline in activity of complexes III and IV by supplementation was significantly associated with cognitive improvement, implicating energy supply as one possible mechanism. These results represent proof of principle that complex dietary supplements can provide powerful benefits for cognitive function and brain aging.
Keywords Cognitive aging, Learning, Aging, Dietary supplements, Mitochondria, Brain mass
From the FULL TEXT Article:
Introduction
Cognitive decline afflicts virtually all aging animals and in humans is also associated with neuropathologies such as Alzheimer’s (AD) and Parkinson’s (PD) diseases (Kausler 1994; Dubois and Pillon 1997; Chen et al. 2001). Modern industrialized societies face rapidly aging cohorts at high risk for cognitive deterioration and pathology. In the USA, AD afflicts 5.3 million (Alzheimer’s Association 2010) and PD ~1 million people (Parkinson’s Disease Foundation, http://www.pdf.org/en/about_pd). Thirteen percent of those older than 65 years of age have AD, and this is the fifth leading cause of death for this cohort (Alzheimer’s Association 2010). By 2050, 115 million people worldwide are predicted to have AD (Ashe and Zahs 2010).
Available treatments amount to several drugs that provide temporary relief or slow progression (Sparks et al. 2005; Kehoe and Wilcock 2007; Relkin et al. 2009). Risk of AD may be reduced, however, by antioxidants (Grundman and Denaney 2002; Holmquist et al. 2007; Cole and Frautschy 2010), healthy diets (Gu et al. 2010), environmental enrichment (Jankowski et al. 2005; Herring et al. 2008), and exercise (Adlard et al. 2005). Preservation of cognitive function into older ages is associated with increased longevity (Barzilai et al. 2006). Maintaining youthful cognitive functions into older ages would yield great benefits for quality of family life and reduce economic and social costs.
Aging humans display progressive loss of spatial navigation skills (Moffat 2009) accentuated in those with even mild AD (Grossi et al. 2007; Hodges 2006; Widmann et al. 2011). Neurobiological and behavioral manifestations of cognitive decline relevant to humans are shared by several animal models (Gallagher and Nicolle 1993) including those genetically engineered to express attributes of human pathologies such AD (e.g., Adlard et al. 2005; Jankowski et al. 2005; Herring et al. 2008). Aging rodents are also especially vulnerable to declining spatial memory (Foster 1999), and this can be quantified by performance in the Morris water maze (MWM) (Brandies et al. 1989; Gallagher and Burwell 1989; Wehner and Silva 1996; Crawley et al. 1997; Adlard et al. 2005; Jankowski et al. 2005). Learning and memory processes tested by the MWM are considered to particularly assess hippocampal function (Crawley et al. 1997; Foster 1999; Adlard et al. 2005).
Age-related accumulation of oxidative damage in brain contributes to cognitive deterioration (Nicolle et al. 2001; Dröge and Schipper 2007; Rollo 2009), but recently, energy metabolism (i.e., age-related decline in mitochondrial function) has also been highlighted (Navarro and Boveris 2007; Aksenov et al. 2010). Mitochondrial complex I dysfunction is strongly implicated in PD (Navarro and Boveris 2009), and complex III is also associated with generation of reactive oxygen species (ROS) relevant to aging (Chen et al. 2004; Muller et al. 2004). Complex IV deficiency was recently identified in AD (Lin and Beal 2006; Luques et al. 2007).

Table 1 Although boosting electron transport chain (ETC) complexes could improve ATP supply, associated increases in mitochondrial respiration could also elevate oxidative stress. Ideally, effective interventions should benefit mitochondrial function while effectively ameliorating ROS damage. We developed a complex dietary supplement (DSP) designed to target five key mechanisms of aging (oxidative stress, inflammation, mitochondrial function, insulin resistance, and membrane integrity) (Table 1). The DSP markedly improved learning in a model of accelerated cognitive aging (transgenic growth hormone mice) (Lemon et al. 2003) and extended longevity of normal mice by 11% (Lemon et al. 2005). It entirely prevented age-related declines in spontaneous motor activity and elevated brain mitochondrial function (complex III) in old age while reducing the rate of free radical generation. Levels of striatal neuropeptide Y were elevated in supplemented normal mice (Aksenov et al. 2010). Additionally, the DSP protected mice from oxidative damage associated with aging and radiation (Lemon et al. 2008a, b; Aksenov et al. 2010).
Previous cognitive benefits of the DSP were demonstrated in transgenic growth hormone mice that express accelerated aging and rapid cognitive decline (Lemon et al. 2003). Here we demonstrate equally striking preservation of cognitive functions in aging normal mice using the MWM. Both supplemented and untreated young mice (<1 year old) learned the water maze well. Untreated animals more than 1 year of age, however, showed no evidence of learning even after 5 days (15 trials). In marked contrast, older supplemented mice showed ~50% better performance than age-matched untreated controls. In fact, learning performance was identical to youth, even for mice ~2 years of age. Although supplemented mice did not differ in size from controls, both genders had significantly greater mass-specific brain mass than untreated mice.
We previously reported a 56% elevation in activity of mitochondrial complex III activity in brains of supplemented mice by ~2 years of age (Aksenov et al. 2010). Similarly, we show here that supplementation achieved a ~65% increase in complex IV activity by ~2 years of age. A correlation analysis suggested a linkage between predicted activity of either mitochondrial complexes III and IV (or their summation) and cognitive performance in aging.
These results represent “proof of principle” that complex nutraceutical formulations can profoundly ameliorate normal cognitive aging. This, combined with our earlier results with transgenic growth hormone mice (that express neuropathology similar to AD) (Lemon et al. 2003), suggests promise for amelioration of human neuropathologies.
Methods
Animals and diets
Breeding and husbandry of mice (random bred C57BL/6J_SJL) were previously described (Lemon et al. 2003). Protocols adhered to the Canada Council on Animal Care. Our DSP contained 30 ingredients (Table 1) available from health food stores without prescription. Daily dosages were derived from recommended human values adjusted for body size and the higher metabolic rate of mice. Supplemented mice received 1 dose/day of DSP that was soaked onto small pieces of bagel in their home cages (details in Lemon et al. 2003; Aksenov et al. 2010). All mice avidly ate the supplement confirming no issues concerning palatability. Mice were randomly assigned at weaning and for life to either the DSP group or untreated controls. Mice (ages 2–31 months) were randomly selected from control and supplemented populations for various experiments.
Morris water maze
Experimental groups comprised 20 untreated and 17 supplemented mice of both genders (ages 3–25 months). Spatial learning was assessed in a circular pool (D 148 cm) filled with water (27 ± 2°C) rendered opaque by adding powdered milk. Room temperature was 23 ± 1°C. The learning task of the mice was to locate a platform (D 11 cm) submerged 1.5 cm below water surface and positioned about 37 cm from the pool wall. An enclosure around the pool provided distal visual cues for spatial orientation. Mice were lowered gently into the water near the wall of the pool, at two alternating starting locations: One position was in the quadrant directly across from the platform (that is, 180° to the platform), and the other position was at 90° to platform. A 120-s time limit was allowed to locate the platform. If unsuccessful, mice were guided to the platform and given 30 s to survey visual cues.
Each mouse was tested over five consecutive days with three trials/day separated by a 40–50-min inter-trial interval. A training schedule with inter-trial intervals of 10–60 min may effectively assess long-term memory acquisition in mice (Kogan et al. 1996). Between-trials mice were dried and placed in cages with a heating pad. Pre-experimental swims (without platform) were conducted prior to the study to reduce emotionality associated with the novelty of swimming and to assess swimming ability (poor swimmers were excluded). Trials were videotaped from above and analyzed using Noldus© EthoVision tracking software to obtain measures of latency to reach the platform and swimming speed.
Brain mitochondrial complex IV activity
We were interested in complex IV activity as it is implicated in Alzheimer’s disease (Lin and Beal 2006; Luques et al. 2007). Complex III was analyzed previously (Aksenov et al. 2010). Normal male mice (n = 18) were used for mitochondrial studies (not tested in the MWM). Brains were removed on ice and stored at ?80°C. For the analysis of complex IV activity (Long et al. 2009), assay buffer contained 50 mmol/l phosphate buffer, 0.1% bovine serum albumin, 0.2% Tween-20, and 60 ?mol/l reduced cytochrome c. Complex IV was measured by monitoring the decrease of reduced cytochrome C at 550 nm.
Brain weight
Brains were collected from a wide age range (2–31 months) of normal supplemented and untreated male and female mice from the breeding colony (n = 182). Mice were decapitated, brains were dissected out, and immediate wet weight was measured. Body mass of each mouse was also recorded, and brain weights were corrected for body mass (e.g., milligrams brain per gram mouse).
Statistical analyses
Effects of diet on learning in young and old mice in the MWM were compared using a t test where latency is reported as mean ± SE. Age-related trends in the levels of complex IV activity were described with linear regressions, and slopes and intercepts were compared using an ANCOVA with age as a covariate. Mass-specific brain weights were characterized with linear regression and impacts of diet were distinguished using t test and ANCOVA. Analyses were performed with Statistica® software.
Results
Morris water maze
Learning in the MWM was evaluated by measuring the latency to locate the escape platform on day 1 (trials 1–3) against day 5 (trials 13–15). Latency is reported separately for young (<1 year old) and old (>1 year old) mice to account for effects of age on learning. Mean ages for young supplemented and untreated mice were 6.2 and 6.7 months and for old supplemented and untreated mice were 19.3 and 19.1 months of age, respectively. No gender differences were observed other than females swam on average ~30% faster than males; however, this was not significantly resolved. Hence, data were pooled for both genders to increase statistical resolution.
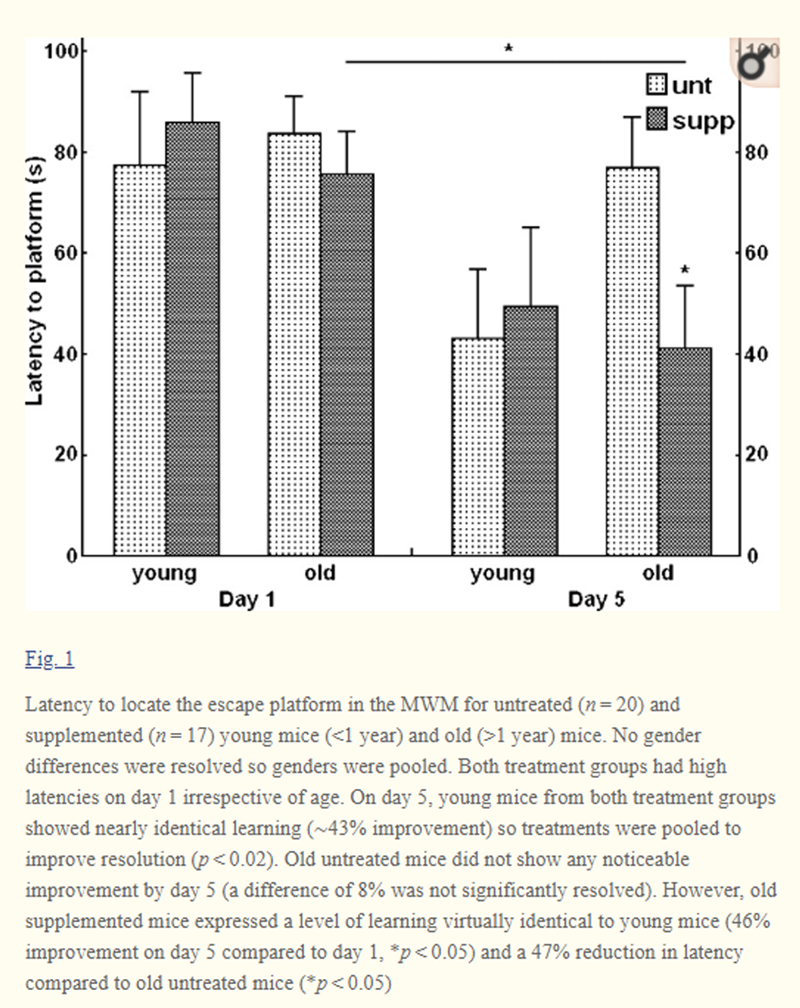
Figure 1 As expected, all mice displayed high latency scores irrespective of age group or dietary supplementation on day 1 (i.e., no significant differences were found between young and old or between supplemented and untreated mice) (Fig. 1). Average latency for all combined groups on day 1 was 81.1 ± 4.5 s. Young mice from both treatments showed a virtually identical decrease in latency on day 5 compared to day 1 (Fig. 1). Since diet did not affect learning in this age group, young mice were pooled to increase statistical resolution. A reduction in latency from 82.2 ± 8.1 s on day 1 to 46.7 ± 10.1 s on day 5 (improvement of 43%) was significantly resolved (p < 0.02) indicating that young mice were successful at learning the maze.
In other studies, some mice employ a search strategy where they circle the pool at a distance from the wall that ensures finding the platform (chaining) (Janus 2004; Patil et al. 2009). With further training, such mice ultimately adopt a direct approach to the platform, indicating spatial learning (Patil et al. 2009). Examination of paths for our mice suggests specific spatial learning rather than chaining.
Contrary to young animals, the DSP had a significant impact on learning of old mice. For old untreated mice, the latency to locate the escape platform remained largely unchanged from day 1 to day 5 (a difference of 8% was not significantly resolved) indicating that the task was not learned (Fig. 1). However, supplemented mice showed a 46% significant decrease in latency from day 1 to day 5 (75.7 ± 8.5 to 41.3 ± 12.5 s; p < 0.05) (Fig. 1). This indicates that old supplemented mice successfully learned the maze, unlike the age-matched untreated counterparts. Additionally, latency to locate the platform for old supplemented mice on day 5 (41.3 ± 12.5 s) was significantly less that of untreated old mice on the same day (77.0 ± 10.0 s) (p < 0.05) reinforcing the finding that learning only occurred in the supplemented group (Fig. 1). Strikingly, day 5 performance of old supplemented mice was virtually identical to that of young animals (41.3 ± 12.5 and 46.7 ± 10.1 s, respectively) (Fig. 1). Hence, it appears that DSP supplementation preserved cognitive function of old mice at youthful levels.
We observed that some animals showed evidence of improvement, but others showed no learning at all (an all-or-nothing pattern). We tested whether the proportion of mice showing no evidence of learning differed between dietary treatments. Mean latency to locate the platform on day 1 (pre-learning trials) was 81 s for the entire population. Therefore, mice with day 5 latency scores greater than or equal to 81 s were considered non-learners since no improvement was seen; all other mice were considered to have learnt the maze to some degree. Based on this modest criterion, ten out of 20 untreated mice (50%) did not learn the maze at all compared to only two out of 17 supplemented mice (12%). Chi-square resolved a significant difference (p < 0.05) between the proportion of non-learners and learners between treatments.
Mitochondrial complex IV activity
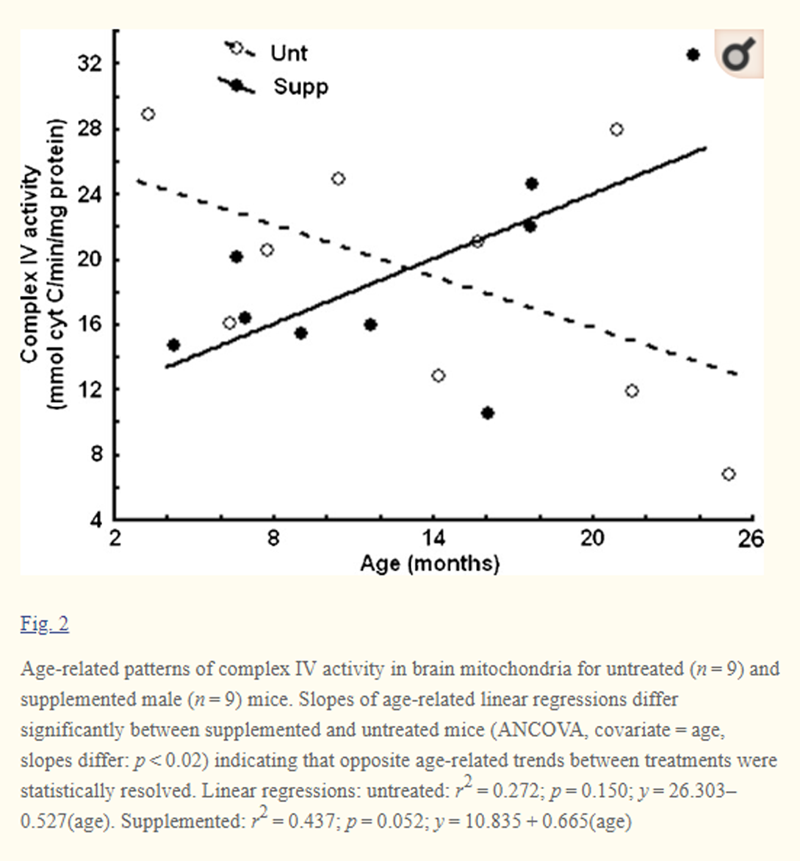
Figure 2 Impact of the DSP on brain mitochondrial complex IV activity was assessed with respect to age. Regression lines relating complex IV activity to age had virtually opposite slopes for untreated (?0.527) versus supplemented mice (+0.665), and differences in slopes and intercepts for these regressions were statistically differentiated (ANCOVA, covariate = age, slopes: p < 0.02, intercepts: p < 0.0001, Fig. 2). Untreated mice had the highest levels of complex IV when young (~100 days/old) but then expressed progressive age-related loss that declined to ~50% that of youthful levels by ~2 years of age (Fig. 2). Conversely, supplemented mice showed a steady age-related rise in complex IV activity and by 2 years of age levels were ~65% higher than in youth (Fig. 2). Thus, the DSP not only prevented age-related decline in brain complex IV activity but it also appears to support the increasing activity with age (Fig. 2).
These findings mirror previous results obtained for complex III activity in brain (Aksenov et al. 2010). For the 18 mice in the current study, the impact of the DSP on complex III activity was more strongly resolved (ANCOVA; p < 0.003) than complex IV (ANCOVA; p < 0.02), but both the magnitude and direction of impacts were very similar. Given the linkage of these complexes and their association with free radical generation, we also considered the impact of the DSP on the summed activities of complexes III and IV. ANCOVA obtained a stronger statistical resolution for the complex III + IV summation than for either individual complex (p < 0.0015).
Brain mitochondrial activity and learning
Table 2 On advice during review, we explored a possible relationship between activity of brain mitochondrial complexes III and IV and performance in the water maze (impacts of DSP on age-related levels of brain complex III activity were previously reported by Aksenov et al. 2010)
. Since measures of mitochondrial activity and learning performance were assessed in different animals, we utilized regression equations to predict mitochondrial complex IV activity for mice of a given age in each treatment group (see Fig. 2). We then examined correlations between predicted values of complex IV activity and learning performance in the MWM (i.e., latency to locate escape platform on day 5). The same procedure was repeated for complex III and for the sum of complexes III + IV. Table 2 shows the correlation between complexes III, IV, and III + IV and learning in the MWM. No correlation was found between mitochondrial complex activity and learning in younger (<1 year old) mice (Table 2). However, for old mice, significant relationships emerged between learning and levels of complexes III, IV, and III + IV activities (p < 0.03, p < 0.05, and p < 0.04; respectively) (Table 2). These results suggest that older mice with higher levels of mitochondrial activity in the brain were better learners.
Brain weight
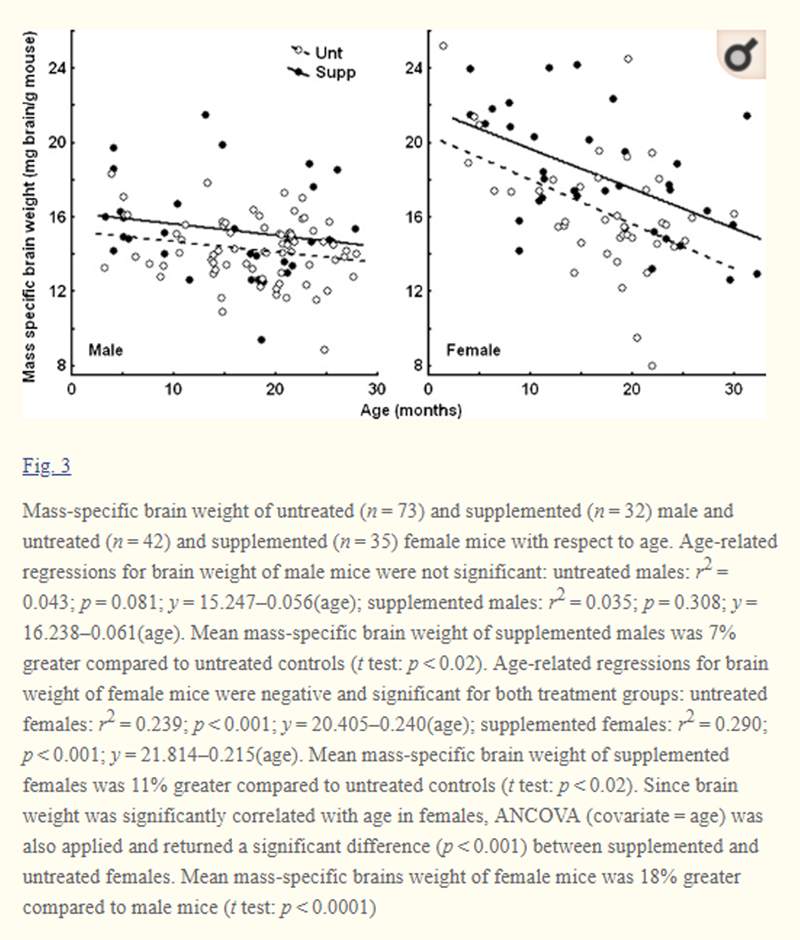
Figure 3 Mass-specific brain weight (i.e., brain weight corrected for body mass) was collected for 115 supplemented mice and 67 untreated mice of both genders. Mean ages for untreated and supplemented males and females were 17.5, 16.3, 16.8, and 16.8 months, respectively. Body mass of females was 13% less than body mass of males (p < 0.00001). However, no difference in body mass was observed between supplemented and untreated mice of either gender, consistent with our previous results (Lemon et al. 2003). Thus, variation in mass-specific brain weights between treatments does not reflect differences in body mass. On average, mass-specific brain weights of females were 18% greater than males (p < 0.0001). Male mice did not show a significant reduction of brain weight with age in either treatment despite apparent trends (Fig. 3). Female mice, however, showed strongly significant reductions in brain weight with age in both treatment groups (p < 0.001) (Fig. 3). Female brain weights were greater in youth, but by ~2 year of age approached those of male mice (Fig. 3). The DSP was associated with significantly larger brains in both male and female mice. Since no significant age-related trends were established in males, a t test was applied. Mean brain weights of supplemented males were 7% greater compared to untreated controls (p < 0.02) (Fig. 3). For females, mean brain weights were 11% grater if supplemented compared to untreated controls (t test: p < 0.02) (Fig. 3). Since weight of female brains was strongly correlated with age, ANCOVA (covariate = age) was also applied confirming that treatment with DSP significantly increased brain weight across all ages (p < 0.001).
Discussion
Morris water maze
The MWM is a standard test of memory and learning for mice and rats and is routinely applied to assess aging and pathology (Crawley et al. 1997; Patil et al. 2009). Strains vary in ability on the MWM, which can relate to variation in anxiety or visual acuity (Klapdor and van der Stay 1996; Crawley et al. 1997). We used C57BL/6J-SJL hybrid mice. One advantage of using hybrid strains is that recessive mutations that might confound interpretation show less penetrance (Lee and Silva 2009). C57BL/6 mice competently learn the MWM but SJL mice (that may express visual impairment) do not (Crawley et al. 1997). Visual acuity of our mice was previously confirmed (Rollo et al. 1999; Lemon et al. 2003), and all mice employed showed intact visual responses.
Our previous results with an eight-choice cued maze documented that transgenic growth hormone mice had remarkably superior learning in youth but by about 1 year of age could not learn the task (Lemon et al. 2003). If supplemented, however, transgenic mice showed no declines but instead learned significantly faster as they aged (Lemon et al. 2003). Thus, the supplement was highly effective in a model of rapid brain deterioration resembling AD. Results presented here importantly attest that the DSL also strongly offsets normal brain aging.
Fifty percent of all untreated mice showed no evidence of learning even after 15 trials over 5 days compared to only two out of 17 mice in the supplemented group (chi-square: p < 0.05). In our previous studies with an eight-choice cued maze, 33% of normal mice (all untreated) also showed no evidence of learning particularly when older. Learning of both tasks tends to be an all-or-none phenomenon, mice either expressing steady incremental improvement or little progress (Lemon et al. 2003; Chaudhry et al. 2008; Rollo et al. 1999). It is all the more remarkable then that older supplemented mice effectively learned the MWM, achieving a 46% reduction in latency to reach the hidden platform by day 5 (Fig. 1). It appears that the DSP does not simply improve cognition but actually enables learning in a large proportion of mice that might otherwise not learn. Moreover, the threshold nature of this learning suggests that higher-order processes associated with “insight” may be favored by the supplement. Cognitive decline is one of the most reliable biomarkers of aging in animals (Carney et al. 1991; Rollo 2009). Thus, prevention of cognitive decline in normal animals spanning ages of 12 to 27 months attests to great efficacy of the supplement (Fig. 1).
We recently documented that our DSP prevented declines in spontaneous motor activity that, like cognition, is regarded as a fundamental biomarker of aging (Aksenov et al. 2010). This raises the possibility that supplementation could change swimming speed which can confound results for the MWM (Klapdor and van der Stay 1996). Mean swimming speed, however, proved virtually identical on either treatment. Given that exercise can benefit cognitive function (Ding et al. 2006; Kramer et al. 2006) and spatial learning (i.e., MWM; Adlard et al. 2005), however, increased spontaneous activity could synergize with the DSP to maintain youthful cognition into advanced ages.
Mitochondrial function
Maintaining effective mitochondrial function was a specific design criterion for our DSP (e.g., α-lipoic acid, acetyl-l-carnitine, B vitamins, coenzyme Q10, maintenance of mitochondrial membranes, and general antioxidant protection). Strong alterations in mitochondrial function indicate that effective dietary components penetrate the blood–brain barrier and impact mitochondria (key criteria sought for anti-aging interventions). We previously reported that untreated mice experienced 46% loss of complex III activity in brain by 2 years of age whereas mice supplemented with the DSP showed a 56% increase (Aksenov et al. 2010). Two-year-old mice had 85% greater complex III activity if supplemented (Aksenov et al. 2010). Here we document a similar age-related pattern for complex IV. Untreated mice showed 40–50% loss of complex IV activity between 3 and 24 months of age (Fig. 2). In contrast, supplemented mice expressed a 65% gain in complex IV activity between 3 and 24 months (Fig. 2). Nutrients altering mitochondrial activity can impact numerous brain functions by modulating ATP availability (Gomez-Pinilla 2008). We know of no other intervention so strongly impacting mitochondrial activity.
Activity of mitochondrial ETC complexes and ATP production generally decline with age (Navarro and Boveris 2007), and declining complex IV activity is a particularly reliable biomarker of aging associated with cognitive loss (Navarro and Boveris 2007). Neurodegenerative conditions (e.g., AD) are also characterized by mitochondrial dysfunction and declining complex IV activity (Albers and Beal 2000; Lin and Beal 2006). Selective reduction of complex IV in rat primary hippocampal neurons results in neuronal death (Atamna et al. 2002). Whether AD is a specific pathology or simply an extreme manifestation of normal senescence remains debatable (Ashe and Zahs 2010). Although previous aging theory highlighted free radical damage as a primary cause, mitochondrial dysfunction and energy shortfalls may contribute to both normal and pathological cognitive declines. Indeed, learning and activity of ETC complexes (III, IV, and III + IV) were positively correlated in mice more than 1 year old (Table 2). Hence, it appears that mitochondrial function is linked to cognitive capacity in old age.
We found no correlation between activity of either complex III or IV and learning of the MWM in young animals (Table 2). Supplemented mice actually showed lower levels of complex IV (Fig. 2) and complex III (Aksenov et al. 2010) activity in youth although this did not compromise cognition (see: Fig. 1) or levels of physical activity (Aksenov et al. 2010).
Brain size
Brain size is positively associated with intelligence in humans (Andreasen et al. 1993), and growth hormone transgenic mice that have larger brains also had vastly superior learning on an eight-choice visually cued maze and novel object recognition and memory tests (Lemon et al. 2003; Chaudhry et al. 2008). Rodents selected for larger brains (and associated increases in body size) also display enhanced learning (Herman and Nagy 1977). Early studies noted great individual differences with respect to aging of human brains (Creasey and Rapoport 1985) suggesting interventions are possible. Reduction in size of most major brain regions occurs in aging resulting in significantly reduced brain mass (Murphy et al. 1992; Allen et al. 2005).
Both our supplemented and untreated mice showed diminishing mass-specific brain weight with age, particularly in females (that also had larger relative brain size than males in youth) (Fig. 3). Supplemented mice maintained larger brains than controls throughout life and in old age brains of supplemented mice were 7% larger in males (p < 0.02) and 11% larger in females (p < 0.02) (Fig. 3). Hippocampal function critical to memory is exceptionally sensitive to aging (Kausler 1994; Apostolova and Thompson 2007). The MWM test is considered to test hippocampal function (Vorhees and Williams 2006). Neurodegeneration of the hippocampus is widely implicated in the loss of spatial memory (navigation skills), dementia, and AD (Foster 1999; Apostolova and Thompson 2007). In AD, the rate of hippocampal atrophy is 3.5–4%/year, exceeding the ~1.6%/year loss observed in normal aging by nearly 3-fold (Jack et al. 1998, 2000).
For normal aging and neurodegenerative conditions alike, diminishing hippocampal volume is correlated with atrophy of other brain regions, increasing ventricular space and diminished brain volume (Scahill et al. 2003; Apostolova and Thompson 2007). Consequently, brain size is a strong predictor of cognitive decline, hippocampal degeneration, and risk of AD (Scahill et al. 2003; Jack et al. 2004). It is likely that the DSP benefited aging hippocampal function.
Complex dietary supplements
A supplement containing 18 vitamins and micronutrients was found to improve cognitive functions in healthy elders other than for long-term memory (Chandra 2001); ten of these are contained in our DSP. Esposito et al. (2002) reviewed literature on antioxidant treatment of various neurodegenerative diseases. Of 13 materials mentioned, our DSP contains eight. In a compilation of nutrients positively impacting cognitive functions, 13 of those identified are present in our DSP (Gomez-Pinilla 2008). A study providing omega 3 fatty acids combined with iron, zinc, folate, and vitamins A, B6, B12, and C to children (five of these are in our DSP) obtained improved scores on tests of verbal intelligence, learning, and memory (Osendarp and the NEMO Study Group 2007). Such results were considered indicative of synergism among ingredients (Gomez-Pinilla 2008). A meta-analysis of studies administering supplements to children that contained at least three ingredients found a marginally significant improvement in fluid intelligence (reasoning) and a significant improvement in academic performance (Eilander et al. 2010).
Zahs and Ashe (2010) compiled a list of interventions that positively impact AD symptoms in APP mouse models. Fourteen of the dietary interventions listed are in our DSP. Human subjects with mild AD showed improvement in the delayed verbal recall task after just 12 weeks on a complex dietary supplement which shares 14 ingredients with our DSP (Scheltens et al. 2010). Of 20 materials considered of possible value in Alzheimer’s in an authoritative review, 12 are in our DSP (Pocernich et al. 2011). Several reviews have suggested that multiple supplements promise to yield the greatest benefits (e.g., Esposito et al. 2002; Pocernich et al. 2011). Our results demonstrate proof of this principle.
Diets high in fruit and vegetables also positively impact brain aging and behavior, but none shows the magnitude of benefits we have documented in this and previous papers (Shukitt-Hale et al. 2008; Joseph et al. 2009). The particular value of specific plants or dietary mixes of fruit and vegetables not only reflect specific content (i.e., flavonoids) bur likely involves interactions and additive effects. Most ingredients in out DSP occur naturally in foods and spices. Unlike normal meals, the DSP combines ingredients specifically targeted to five mechanisms of aging and provides these in relatively concentrated form
Conclusion
Despite generally disappointing results in studies applying one or a few dietary supplements, our results show that complex supplements can powerfully impact brain aging in normal animals as well as neuropathology in a transgenic mouse. The DSP had a relatively broad spectrum of actions: It augmented mitochondrial function, increased mass-specific brain weight, and markedly improved cognitive abilities in old age. Our previous studies demonstrated that it elevated expression of brain neurotransmitter systems and ameliorated age-related motor decline (Aksenov et al. 2010). Such effects likely emerge from additive, synergistic, and recycling interactions among ingredients and the fact that the DSP was designed to ameliorate multiple mechanisms of aging. Since materials in DSP are available in health food stores, a large sector of the aging human population could obtain nearly immediate access should human trials confirm safety and benefits.
Acknowledgments
This research was supported by a grant to CDR from the Natural Sciences and Engineering Research Council of Canada. We thank Zoya Tov for her contributions to diet preparation and technical support for the experiments.
REFERENCES
Adlard PA, Perreau VM, Pop V, Cotman CW.
Voluntary exercise decreases amyloid load in transgenic model of Alzheimer’s disease.
J Neurosci. 2005;25:4217–4221Aksenov V, Long J, Lokuge S, Foster JA, Liu J, Rollo CD.
A dietary supplement ameliorates locomotor, neurotransmitter and mitochondrial aging.
Exp Biol Med. 2010;335:66–76Albers DS, Beal MF.
Mitochondrial dysfunction and oxidative stress in aging and neurodegenerative disease.
J Neural Transm Suppl. 2000;59:133–154Allen JS, Bruss J, Brown CK, Damasio H.
Normal neuroanatomical variation due to age: the major lobes and a parcellation of the temporal region.
Neurobiol Aging. 2005;26:1245–1260Alzheimer’s Association Alzheimer’s disease facts and figures.
Alzheimers Dement. 2010;6:1–70Andreasen NC, Flaum M, Swayze V, O’Leary DS, Alliger R, Cohen G, Ehrhardt J, Yuh NT.
Intelligence and brain structure in normal individuals.
Am J Psychiatry. 1993;150:130–134Apostolova LG, Thompson PM.
Brain mapping as a tool to study neurodegeneration.
Neurotherapeutics. 2007;4:387–40059Ashe KH, Zahs KR.
Probing the biology of Alzheimer’s disease in mice.
Neuron. 2010;66:631–645Atamna H, Killilea DW, Killilea AN, Ames BN.
Heme deficiency may be a factor in the mitochondrial and neuronal decay of aging.
PNAS. 2002;99:14807–14812Barzilai N, Atzmon G, Derby CA, Bauman JM, Lipton RB.
A genotype of exceptional longevity is associated with preservation of cognitive function.
Neurology. 2006;67:2170–2175Brandies R, Brandies Y, Yehuda S.
The use of the Morris water maze in the study of memory and learning.
Int J Neurosci. 1989;48:29–69Carney JM, Starke-Reed PE, Oliver CN, Landum RW, Cheng MS, Wu JF, Floyd RA.
Reversal of age-related increase in brain protein oxidation, decrease in enzyme activity, and loss in temporal and spatial memory by chronic administration of the spin-trapping compound N-tert-butyl- α-phenylnitrone.
PNAS. 1991;88:3633–3636Chandra RK.
Effect of vitamin and trace-element supplementation on cognitive function in elderly subjects.
Nutrition. 2001;17:709–712Chaudhry AM, Marsh-Rollo SE, Aksenov V, Rollo CD, Szechtman H.
Modifier selection by transgenes: the case of growth hormone transgenesis and hyperactive circling mice.
Evol Biol. 2008;35:267–286Chen P, Ratcliff G, Belle SH, Cauley JA, DeKosky ST, Ganguli M.
Patterns of cognitive decline in presymptomatic Alzheimer disease.
Arch Gen Psychiatry. 2001;58:853–858Chen Q, Vazquez EJ, Moghaddas S, Hoppel CL, Lesnefsky EJ.
Production of reactive oxygen species by mitochondria: central role of complex III.
J Biol Chem. 2004;278:36027–36031Cole GM, Frautschy SA.
DHA may prevent age-related dementia.
J Nutr. 2010;140:869–874Crawley JN, Belknap JK, Collins A, et al.
Behavioral phenotypes of inbred mouse strains: implications and recommendations for molecular studies.
Psychopharmacology. 1997;132:107–124Creasey H, Rapoport SI.
The aging human brain.
Ann Neurol. 1985;17:2–10Ding Q, Vaynman S, Akhavan M, Ying Z, Gomez-Pinilla F.
Insulin-like growth factor I interfaces with brain-derived neurotrophic factor-mediated synaptic plasticity to modulate aspects of exercise-induced cognitive function.
Neuroscience. 2006;140:823–833Dröge W, Schipper HM.
Oxidative stress and aberrant signaling in aging and cognitive decline.
Aging Cell. 2007;6:361–370Dubois B, Pillon B.
Cognitive deficits in Parkinson’s disease.
J Neurol. 1997;244:2–8Eilander A, Gera T, Sachdev HS, Transler C, van der Knaap HC, Kok FJ, Osendarp SJ.
Multiple micronutrient supplementation for improving cognitive performance in children: systematic review of randomized controlled trials.
Am J Clin Nutr. 2010;91:115–130Esposito E, Rotilio D, Di Matteo V, Di Giulio C, Cacchio M, Algeri A.
A review of specific dietary antioxidants and the effects on biochemical mechanisms related to neurodegenerative processes.
Neurobiol Aging. 2002;23:719–735Foster TC.
Involvement of hippocampal synaptic plasticity in age-related memory decline.
Brain Res Rev. 1999;30:236–249Gallagher M, Burwell RD.
Relationship of age-related decline across several behavioral domains.
Neurobiol Aging. 1989;10:691–708Gallagher M, Nicolle M.
Animal models of normal aging: relationship between cognitive decline and markers in hippocampal circuitry.
Behav Brain Res. 1993;57:155–162Gomez-Pinilla F.
Brain foods: the effects of nutrients on brain function.
Nat Rev Neurosci. 2008;9:568–578Grossi D, Fasanaro AM, Cecere R, Salzano S, Trojano L.
Progressive topographical disorientation: a case of focal Alzheimer’s disease.
Neurol Sci. 2007;28:107–110Grundman M, Denaney P.
Antioxidant strategies for Alzheimer’s disease.
Proc Nutr Soc. 2002;61:191–202Gu Y, Nieves JW, Stern Y, Luchsinger JA, Scarmeas N.
Diet and prevention of Alzheimer disease.
JAMA. 2010;303:2519–2520Herman BH, Nagy ZM.
Development of learning and memory in mice genetically selected for differences in brain weight.
Dev Psychol. 1977;10:65–75Herring A, Yasin H, Ambrée O, Sachser N, Paulus W, Keyvani K.
Environmental enrichment counteracts Alzheimer’s neurovascular dysfunction in TgCRND8 mice.
Brain Pathol. 2008;18:32–39Hodges JR.
Alzheimer’s centennial legacy: origins, landmarks and the current status of knowledge concerning cognitive aspects.
Brain. 2006;129:2811–2822Holmquist L, Stuchbury G, Berbaum K, et al.
Lipoic acid as a novel treatment for Alzheimer’s disease and related dementias.
Pharmacol Ther. 2007;113:154–164Jack CR, Jr, Petersen RC, Xu Y, et al.
Rate of medial temporal lobe atrophy in typical aging and Alzheimer’s disease.
Neurology. 1998;51:993–999Jack CR, Jr, Petersen RC, Xu Y, et al.
Rates of hippocampal atrophy correlate with change in clinical status in aging and AD.
Neurology. 2000;55:484–489Jack CR, Jr, Shiung MM, Gunter JL, et al.
Comparison of different MRI brain atrophy rate measures with clinical disease progression in AD.
Neurology. 2004;62:591–600Jankowski JL, Melnikova T, Fadale DJ, et al.
Environmental enrichment mitigates cognitive deficits in a mouse model of Alzheimer’s disease.
J Neurosci. 2005;25:5217–5224Janus C.
Search strategies used by APP transgenic mice during navigation in the Morris water maze.
Learn Mem. 2004;11:337–346Joseph AJ, Shukitt-Hale B, Willis LM.
Grape juice, berries and walnuts affect brain aging and behavior.
J Nutr. 2009;139:1813S–1817SKausler DH.
Learning and memory in normal aging. San Diego: Academic; 1994.Kehoe PG, Wilcock GK.
Is inhibition of the renin–angiotensin system a new treatment option for Alzheimer’s disease?
Lancet Neurol. 2007;6:373–378Klapdor K, van der Stay FJ.
The Morris water-escape task in mice: strain differences and effects of intra-maze contrast and brightness.
Physiol Behav. 1996;60:1247–1254Kogan JH, Frankland PW, Blendy JA, Coblentz J, Marowitz Z, Schütz G, Silva AJ.
Spaced training induces normal long-term memory in CREB mutant mice.
Curr Biol. 1996;7:1–11Kramer AF, Erickson KI, Colcombe SJ.
Exercise, cognition and the aging brain.
J Appl Physiol. 2006;101:1237–1242Lee YS, Silva AJ.
The molecular and cellular biology of enhanced cognition.
Nat Rev Neurosci. 2009;10:126–140Lemon JA, Boreham DR, Rollo CD.
A dietary supplement abolishes age-related cognitive decline in transgenic mice expressing elevated free radical processes.
Exp Biol Med. 2003;228:800–810Lemon JA, Boreham DR, Rollo CD.
A Complex Dietary Supplement Extends Longevity of Mice
J Gerontol A Biol Sci Med Sci. 2005 (Mar); 60 (3): 275–279Lemon JA, Rollo CD, Boreham DR.
Elevated DNA Damage in a Mouse Model of Oxidative Stress: Impacts of Ionizing Radiation and a Protective Dietary Supplement
Mutagenesis. 2008 (Nov); 23 (6): 473–482Lemon JA, Rollo CD, McFarlane NM, Boreham DR.
Radiation-induced apoptosis in mouse lymphocytes is modified by a complex dietary supplement: the effect of genotype and gender.
Mutagenesis. 2008;23:465–472Lin MT, Beal MF.
Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases.
Nature. 2006;443:787–795Long J, Gao F, Tong L, Cotman CW, Ames BN, Liu J.
Mitochondrial decay in the brains of old rats: ameliorating effect of alpha-lipoic acid and acetyl-L-carnitine.
Neurochem Res. 2009;34:755–763Luques L, Shoham S, Weinstock M.
Chronic brain cytochrome oxidase inhibition selectively alters hippocampal cholinergic innervation and impairs memory: prevention by ladostigil.
Exp Neurol. 2007;206:209–219Moffat SD.
Aging and spatial navigation: what do we know and where do we go?
Neuropsychol Rev. 2009;19:478–489Muller FL, Liu Y, Van Remmen H.
Complex III releases superoxide to both sides of the inner mitochondrial membrane.
J Biol Chem. 2004;279:49064–49073Murphy DGM, DeCarli C, Schapiro MB, Rapoport SI, Horwitz B.
Age-related differences in volumes of subcortical nuclei, brain matter, and cerebrospinal fluid in healthy men as measured with magnetic resonance imaging.
Arch Neurol. 1992;49:839–845Navarro A, Boveris A.
The mitochondrial energy transduction system and the aging process.
Am J Physiol Cell Physiol. 2007;292:C670–C686Navarro A, Boveris A.
Brain mitochondrial dysfunction and oxidative damage in Parkinson’s disease.
J Bioenerg Biomembr. 2009;41:517–521Nicolle MM, Gonzalez J, Sugaya K, et al.
Signatures of hippocampal oxidative stress in aged spatial learning-impaired rodents.
Neuroscience. 2001;107:415–431Osendarp SJ,
the NEMO Study Group Effect of a 12-mo micronutrient intervention on learning and memory in well-nourished and marginally nourished school-aged children: 2 parallel, randomized, placebo-controlled studies in Australia and Indonesia.
Am J Clin Nutr. 2007;86:1082–1093Patil SS, Sunyer B, Hoger H, Lubec G. Evaluation of spatial memory of C57BL/6J and CD1 mice in the Barnes maze, the multiple T-maze and in the Mor
ris water maze.
Behav Brain Res. 2009;198:58–68Pocernich CB, Bader Lange ML, Sultana R, Butterfield DA.
Nutritional approaches to modulate oxidative stress in Alzheimer’s disease.
Curr Alzh Res. 2011;8:452–469Relkin NR, Szabo P, Adamiac B, et al.
18-month study of intravenous immunoglobulin for treatment of mild Alzheimer’s disease.
Neurobiol Aging. 2009;30:1728–1736Rollo CD.
Dopamine and aging: intersecting facets.
Neurochem Res. 2009;34:601–629Rollo CD, Ko CV, Tyerman JGA, Kajiura L.
The growth hormone axis and cognition: empirical results and integrated theory derived from giant transgenic mice.
Can J Zool. 1999;77:1874–1890Scahill RI, Frost C, Jenkins R, et al.
A longitudinal study of brain volume changes in normal aging using serial registered magnetic resonance imaging.
Arch Neurol. 2003;60:989–994Scheltens P, Kamphuis PJ, Verhey FR, et al.
Efficacy of a medical food in mild Alzheimer’s disease: a randomized, controlled trial.
Alzheimers Dement. 2010;6:1–10e.1Shukitt-Hale B, Lau FC, Joseph JA.
Berry fruit supplementation and the aging brain.
J Agric Food Chem. 2008;56:636–641Sparks DL, Sabbagh MN, Connor DJ, et al.
Atorvastatin for the treatment of mild to moderate Alzheimer’s disease.
Arch Neurol. 2005;62:753–757Vorhees CV, Williams MT.
Morris water maze: procedures for assessing spatial and related forms of learning and memory.
Nat Protoc. 2006;1:848–858Wehner JM, Silva A.
Importance of strain differences in evaluations of learning and memory processes in null mutants.
MRDD Res Rev. 1996;2:243–248Widmann CN, Beinhoff U, Riepe MW (2011)
Everyday memory deficits in very mild Alzheimer’s disease.
Neurobiol AgingZahs KR, Ashe KH.
‘Too much good news’—are Alzheimer mouse models trying to tell us how to prevent, not cure, Alzheimer’s disease?
Trends Neurosci. 2010;33:381–389
Return to NUTRITION
Since 8–07-2018
| Home Page | Visit Our Sponsors | Become a Sponsor |
Please read our DISCLAIMER |