

Nutrient Biomarker Patterns, Cognitive Function,
and MRI Measures of Brain AgingThis section is compiled by Frank M. Painter, D.C.
Send all comments or additions to: Frankp@chiro.org


FROM: Neurology. 2012 (Jan 24); 78 (4): 241–249 ~ FULL TEXT
Bowman GL, Silbert LC, Howieson D, Dodge HH, Traber MG,
Frei B, Kaye JA, Shannon J, Quinn JF.
Department of Neurology,
Oregon Health & Science University,
Portland, USA.
bowmang@ohsu.edu
Nutrients That Strengthen Older Brains
Higher blood levels of omega-3 fatty acids, vitamin B, vitamin C, vitamin D and vitamin E are associated with better mental functioning in the elderly, a new study has found.
OBJECTIVE: To examine the cross-sectional relationship between nutrient status and psychometric and imaging indices of brain health in dementia-free elders.
METHODS: Thirty plasma biomarkers of diet were assayed in the Oregon Brain Aging Study cohort (n = 104). Principal component analysis constructed nutrient biomarker patterns (NBPs) and regression models assessed the relationship of these with cognitive and MRI outcomes.
RESULTS: Mean age was 87 ± 10 years and 62% of subjects were female. Two NBPs associated with more favorable cognitive and MRI measures: one high in plasma vitamins B (B1, B2, B6, folate, and B12), C, D, and E, and another high in plasma marine omega-3 fatty acids. A third pattern characterized by high trans fat was associated with less favorable cognitive function and less total cerebral brain volume. Depression attenuated the relationship between the marine omega-3 pattern and white matter hyperintensity volume.
CONCLUSION: Distinct nutrient biomarker patterns detected in plasma are interpretable and account for a significant degree of variance in both cognitive function and brain volume. Objective and multivariate approaches to the study of nutrition in brain health warrant further study. These findings should be confirmed in a separate population.
From the Full-Text Article:
Introduction
The epidemiology of Alzheimer disease (AD) suggests a role for nutrition. [1–7] Despite studies in favor of a single or a few nutrients in the prevention of AD, the translation to formal clinical trials testing vitamin E, B vitamins, or docosahexaenoic acid have been disappointing. [8–12] Given the interactive nature of nutrient action and metabolism, it is not surprising that a single or few nutrient approaches for neurodegenerative disease are tenuous. [13–15] These results impart the rationale for novel methodologic approaches that appreciate the interactive features of nutrients and model their collective influence in the promotion of brain health.
Food frequency questionnaires (FFQ) have traditionally been used to construct dietary patterns. [16] FFQ is relatively inexpensive and fairly comprehensive, but this method is subject to faulty recall of dietary intake and does not account for variability in nutrient absorption, both of which are issues in the elderly. [17, 18] We have recently reported a reliable blood test that assesses nutritional status in people at risk for dementia. [19] In the current study, we examine the relationship of nutrient biomarkers with cognitive function and MRI.
To capture the effect of nutrients in combination, we construct nutrient biomarker patterns using principal component analysis (PCA). Cluster analysis, [20] index scores, [21] and reduced rank regression [22] have each been applied to FFQ data to assemble dietary patterns, but none have applied PCA to biological markers of diet. One goal is to define dietary patterns that promote cognitive health in the same manner that dietary approaches for hypertension have been derived and applied. [23]
METHODS
Population
The Oregon Brain Aging Study (OBAS) was initiated in 1989 and recruited 293 community-dwelling men and women aged 65 years and older who were generally free of usual confounding factors known to modify risk for cognitive decline (i.e., vascular disease, hypertension, diabetes) to permit a purer study of aging effects on brain parameters. [e1, e2] The current cohort density is 76 and assessments are performed until death. The MMSE, neuropsychological assessments, and the Clinical Dementia Rating (CDR) reflect cognitive status. Participants attend annual study visits with a collateral historian for an evaluation by a staff neurologist, neuropsychologist, and research study member for clinical and cognitive evaluation, MRI, and blood collection. This study utilized circa 2006–2007 banked specimens and participants with a CDR >0.5 were excluded. Plasma, clinical, and neuropsychological data were available for 104 subjects and 42 had MRI within a month of the blood draw.
Standard protocol approval and patient consent
Informed consent was obtained from all patients for participation in this study, which was approved by the OHSU institutional review board for human study.
Nutrient biomarker acquisition and analysis
Preferably, fasting plasma was collected between 07:00 and 12:00 noon Pacific Time beginning in September 2006 and ending December 2007. Heparin plasma was deproteinized with 10% metaphosphoric acid and analyzed for ascorbic acid using high-performance liquid chromatography (HPLC). [e3] Ethylenediaminetetraacetic acid (EDTA) plasma carotenoids, tocopherol, and retinol were analyzed by HPLC using diode array detector and fluorescence detection. [e4] EDTA plasma thiamin, riboflavin, niacin, and pyridoxal 5-phoshate were analyzed by liquid chromatography–mass spectrometry/mass spectrometry. [e4] EDTA plasma folate and vitamin B12 was measured with a chemiluminescence-based assay on an Immulite analyzer (Siemens Corporation, Washington, DC). Radioimmunoassay measured EDTA plasma 25-OH vitamin D (Immunodiagnostics Systems Inc., Scottsdale, AZ). Gas chromatography equipped with a flame ionization detector quantified plasma fatty acid concentrations. [e4] Plasma lipids were measured with standard enzymatic methods. Reliability statistics for these assays are readily available. [19]
Neuropsychological tests
The battery includes the following: Trail Making Test, Consortium to Establish a Registry for Alzheimer's Disease Word List acquisition and delayed recall, abbreviated Boston Naming Test, Wechsler Memory Scale–Revised Logical Memory Story A I and II and Wechsler Adult Intelligence Scale–Revised Digit Span and Block Design, and the abbreviated Geriatric Depression Scale.
Volumetric MRI acquisition and analysis
Regional volumes of interest are scaled using a 1.5-T MRI and semiautomated REGION image analysis software. Recursive regression analysis of bifeature space based on relative tissue intensities was used to separate tissue types on each coronal image. The sums of pixel areas for all slices were converted to volumetric measures by multiplying by the slice thickness for each of the following regions of interest: total white matter hyperintensity volume (WMH, includes periventricular and subcortical deep signals) and supratentorial brain volume as total cerebral brain volume (TCBV, excluding cerebellum and brainstem). Regression for brain tissue, CSF, and WMH collectively against bone creates a boundary along the inner table of the skull to determine the total intracranial volume (TIV). Additional boundaries were manually traced along the tentorium cerebelli and the superior border of the superior colliculus, the pons, and the fourth ventricle. The pituitary, vessels in the sphenoid region, and any sinuses that may have been included by the automatic regression were excluded manually. All REGION methods have an intraclass correlation coefficient (ICC) of 0.95 or better except WMH volume (ICC = 0.85).
Covariates
PCR assay determined APOE4 carrier status. [e5] Age, gender, years of education, body mass index, socioeconomic status, [e6] blood pressure, current drinking and smoking, active diabetes and hypertension, active depression within the past 2 years, Hachinski scale, [e7] current multivitamin use, duration of fasting, and plasma creatinine were queried from clinician interviews.
Statistical analysis
Descriptive statistics were calculated for demographic, clinical, and nutritional measures.Nutrient biomarker pattern construction. Eight distinct nutrient biomarker patterns (NBPs) were extracted from the original set of 30 biomarkers via multivariate analysis (PCA). [e8, e9] An eigenvalue of 1.0 was set a priori to determine the NBPs to carry into hypothesis testing. Each participant receives a standardized NBP score for each pattern that corresponds to a linear combination of the plasma nutrients that load heavily within each pattern.
Cognitive z scores and MRI. Cognitive z scores generated on the study sample (n = 104) were conceptually combined to represent specific cognitive domains [e10] and combined all together to generate a global cognitive z score (table e-1 on the Neurology® Web site at www.neurology.org). MRI of the supratentorial brain divided by intracranial volume adjusted for differences in head size and the sum of the periventricular and subcortical deep white matter signals divided by the supratentorial volume adjusted for brain size prior to hypothesis testing.
Primary models. The outcomes of interest include the following: cognitive z scores (global and domain specific), TCBV (% of intracranial volume), and WMH volume (% of TCBV). Potential confounders considered for inclusion were based on a previously recognized relationship with the outcomes and an association identified with the NBPs in this study. Two linear regression models were fit for each cognitive and MRI outcome: model 1 includes all 8 NBPs entered simultaneously plus age, gender, education, and APOE4, and model 2 is further adjusted for hypertension and depression simultaneously.
Exploratory analysis. To appreciate the proportion of variance explained in the brain outcomes by the NBPs, we examined the R2 generated by the clinical-demographic variables initially and again with all the NBPs included in the model simultaneously. To explore the hypothesis that WMH and brain atrophy mediate the association between NBPs and cognitive function, we examined changes in significance and coefficient magnitude with and without MRI measures in the model. All analyses were performed with IBM SPSS statistical software, version 19 for Macintosh. All p values reported are 2-sided.
RESULTS
Demographic, clinical, and nutritional characteristics

Table 1 As expected, comorbidities and vascular risk factors were low, with the exception of hypertension (44%). The study cohort is 62% female and 10% carry the APOE4 allele. The mean MMSE was 27 and no participants had a CDR >0.5 (Table 1). The overall nutritional status in the sample population was mostly replete with prevalence of vitamin B12 deficiency (<200 pg/mL) at 7% and vitamin D deficiency (<20 ng/mL) at 25% (table e-2). Participants with MRI (n = 42) were 85 years and older (mean age = 92.6, SD 3.8, range 85–101). Mean age difference between those with and without MRI was 9.4 years (p < 0.0001). The other demographic and clinical characteristics were not different (data not shown).
Nutrient biomarker pattern construction and interpretation
Table 2 Table 2 displays the composition of 8 NBPs. After the eighth NBP extraction with PCA, 72.8% of the total variance was accounted for in the original set of nutrient biomarkers (Table 2, cumulative % variance after NBP8 extraction). For the sake of reference in the following results, NBP1 is described as the BCDE pattern (all loading coefficients > 0.50). NBP2 is described as the saturated fat pattern, NBP3 as the carotenoid pattern, NBP4 as the cholesterol pattern, NBP5 as the marine ω-3 fatty acid pattern, NBP6 as the ω--6 + retinol pattern, NBP7 as the lutein + high-density lipoprotein (HDL) cholesterol pattern, and NBP8 as the trans fat pattern mostly represented by trans linolelaidic acid (18:2ω-6t).
Nutrient biomarker patterns and demographic–clinical characteristics
The NBP1-BCDE and NBP5-marine ω-3 patterns were not associated with any demographic–clinical characteristics. The mean NBP8–trans fat score was 0.713 SD units higher in subjects with hypertension. These findings and the remaining for the other NBPs are presented in table e-3.
Nutrient biomarker patterns and cognitive function
Table 3 The NBP1-BCDE and NBP8–trans fat patterns were the most significant to cognitive function (Table 3). Subjects with higher BCDE scores had better global cognitive function, particularly in domains of executive, attention, and visuospatial function. Participants with higher plasma trans fat scores had worse cognitive function overall (memory, attention, language, processing speed, and global). Subjects with higher NBP5-marine ω-3 scores had better executive function. Memory was better in those with higher NBP7-lutein + HDL cholesterol scores. Memory and language were worse in those with higher NBP6-ω-6 + retinol scores. Adjustment for age, gender, education, APOE4, hypertension, and depression did not attenuate these relationships. Each 1-SD increase in BCDE score associated with a 0.28-SD increase in global cognitive score. Each 1-SD increase in the trans fat score associated with a 0.30-SD decrease in global cognitive score.
Nutrient biomarker patterns and MRI
Table 4
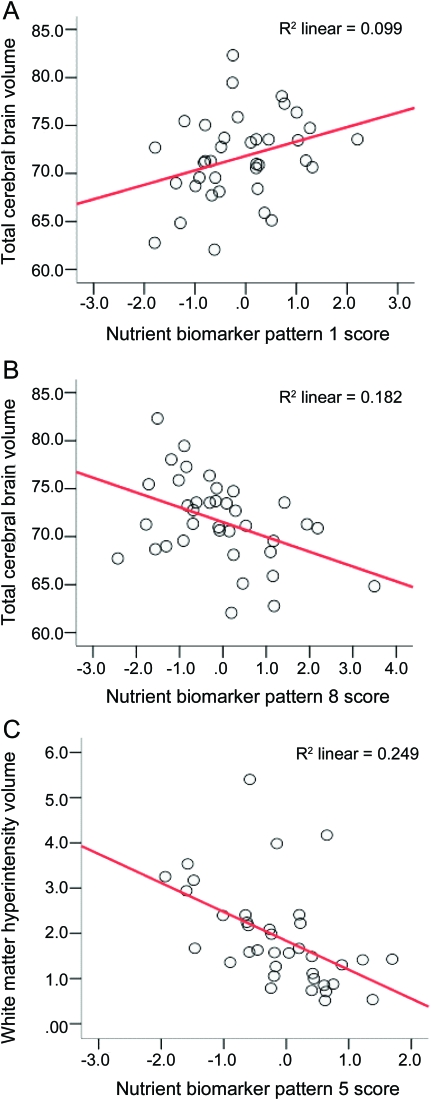
Figure Subjects with higher plasma BCDE scores had more TCBV and those with higher trans fat scores had less TCBV. Subjects with higher marine ω-3 scores had less WMH volume, but after adjustment for depression and hypertension the association was attenuated (Table 4, WMH model 2). Significance of the ω-3 to WMH was lost after adding depression to model 1 (p = 0.030 to 0.097). Adding hypertension had no effect. After stratifying by depression in model 1 for WMH it was apparent that ω-3s were significant only in those without depression (ß = -0.845, p = 0.021). The unadjusted proportions of variance explained in brain volumes by each significant NBP are provided in the Figure.
Exploratory analysis.
Age, gender, education years, APOE4 carrier status, depression, and hypertension together explained 46% of the variation in the global cognitive z score. Adding the NBPs explained an additional 17% (global cognitive z score: R2Covariates = 0.46; R2Covariates + NBPs = 0.63). In regards to the MRI-TCBV, the covariates explained 40% of the total variation and the NBPs explained an additional 37% (TCBV: R2Covariates = 0.396; R2Covariates + NBPs = 0.766). The covariates explained 52% of the WMH variation and the NBPs explained an additional 9% (WMH: R2covariates = 0.512, R2covariates + NBPs = 0.611).
To explore the hypothesis that the NBP effects on cognition are mediated through brain atrophy and WMH, we examined the changes to the association after adding MRI measures to the model. In the reduced subset with MRI available (n = 42), the association between NBP8–trans fat scores and global cognitive function was undetectable (p = 0.054), leaving us unable to pursue this hypothesis for trans fat. However, the relationship between NBP1-BCDE scores and global function was maintained in this subset, and adding MRI measures did not attenuate this association (ß = 0.614, p = 0.010).
DISCUSSION
This cross-sectional study describes the nutrient biomarker patterns identified in plasma from a sample of elders at risk for dementia. This objective and multivariate approach yielded 3 distinct NBPs significant to both cognitive function and MRI measures of brain aging. To our knowledge, this is the first study to apply principal components analysis to biological markers of diet.
Dietary patterns associated with cognitive decline or Alzheimer incidence have historically derived the patterns from FFQ data. Dietary intake can be indexed as “healthy” or “unhealthy” based on existing knowledge and examined in relation to disease risk. [21, 24] Data-driven cluster analysis places subjects into exclusive dietary patterns a posteriori [20] and reduced rank regression combines existing knowledge and the data at hand to derive dietary patterns. [22] These studies using FFQ have identified an intake higher in dark and green leafy vegetables, cruciferous vegetables, [22] fish, [25] and fruit [21, 22] and lower in organ meats, red meat, high-fat dairy, butter, [22] and trans fat [26] as favorable for cognitive health. In thinking about the plasma signature of this diet, we propose that the favorable BCDE pattern and ω-3 pattern would be sensitive to the frequent consumption of dark and green leafy and cruciferous vegetables, fruit, and fish. In addition, a NBP high in trans fat and retinol would be expected in people frequently consuming bakery and fried foods, margarine spreads, [27] red meat, [27] and offal. [28] These consistencies are encouraging and provide impetus for further development of biological markers of diet.
The neuroimaging results suggest that the mechanisms through which the 2 favorable patterns (NBP1-BCDE and NBP5-marine ω-3) affect cognitive function are distinct. Cognitive benefit gained by a plasma profile high in antioxidants C and E, B vitamins, and vitamin D may partially operate on the neurobiology that governs rate of total brain atrophy (e.g., Alzheimer type pathology), whereas the effects of the marine ω-3s may be mediated through more vascular mechanisms. [29, 30] The favorable relationship between the BCDE pattern and global cognitive function was maintained after adding TCBV to the model in our study. This suggests that the effects of this combination on cognition are not entirely mediated through structural changes. Other mechanisms through which this pattern may offer cognitive benefit include the promotion of hippocampal neurogenesis, [31] reduction of ß-secretase activity, [32] oxidative stress, [33, 34] and hyperhomocysteinemia-induced neurotoxicity, [35] and perhaps by maintaining blood–brain barrier integrity. [36]
The high trans fat pattern was consistently associated with worse cognitive performance and less TCBV. Linolelaidic acid is predominantly found in bakery foods such as cookies, doughnuts, cakes, pastries, and pies. [27] These foods are often prepared with hydrogenated vegetable oils to allow for a long shelf life. Higher trans fatty acid intake increases cardiovascular risk, systemic inflammation, and endothelial dysfunction, all of which may explain an association with cognition. [37, 38] Unfortunately, very few studies have assessed trans fat and risk for cognitive decline. [26] Trans fat may aggravate cognitive function independently and jointly through interaction with other dietary factors. [e11] Trans fat may displace DHA in neuronal membranes, but apparently does not impact the neuropathologic Alzheimer hallmarks in mice. [39] The consistency of the association of plasma trans fat with poorer cognitive function and more brain atrophy suggests neurologic consequences in humans, but these findings need to be confirmed.
PCA of fatty acids expressed as weight percentages of total in serum and in erythrocyte membranes have been studied. [e12, e13] The patterns, including eicosapentaenoic and docosahexaenoic acid loading together, were similar to our findings using fatty acids expressed as absolute concentrations in plasma. The interactive metabolism of EPA and DHA, in addition to the similar dietary sources, may explain why these 2 fatty acids load together. PCA constructs the patterns on a basis of collinearity, and this “relatedness” may be partially attributed to interactive metabolism when applied to biological markers of diet. Our observation that the carotenoids (NBP3), total and low-density lipoprotein cholesterol (NBP4), saturated fats (NBP2), and the ω-6 fatty acids (NBP6) load together adds further support to the notion that interactive metabolism is a contributor to NBP construction.
There are limitations of this study. PCA may require investigator decisions with the data in hand. For example, using an eigenvalue of >1.0 as inclusion criteria for the number of patterns extracted to carry forward into hypothesis testing may require more field-specific criteria. Our nutrient biomarkers were selected a priori capitalizing on existing knowledge of an association with neurodegeneration, but this may not reflect the ideal set. Observational studies are susceptible to residual confounding, and our cross-sectional design is not suited for inferring any causal association since the temporal relationship is unattainable. Our sample population was restricted to a relatively healthy and well-educated cohort of white, non-Hispanic elders with minimal genetic risk for AD. These attributes may limit the generalizability of the results.
Future studies should consider validating the external consistency of these findings. The ability of NBPs to predict cognitive and brain volume changes would offer more compelling data. Gene–nutrient interactions underlying a relationship between nutrition and cognition may be important to consider since APOE4 carriers may benefit less from nutritional interventions. [6, 10, 40] The significance of these NBPs at different stages of cognitive status are unknown. These studies will decipher the key nutrient combinations and the population best suited for intervention studies.

Return to B COMPLEX
Return to ALZHEIMER's
Return to OMEGA-3 FATTY ACIDS
Return to NEURODEGENERATIVE DISEASES & EFAs
Since 1-03-2012
| Home Page | Visit Our Sponsors | Become a Sponsor |
Please read our DISCLAIMER |